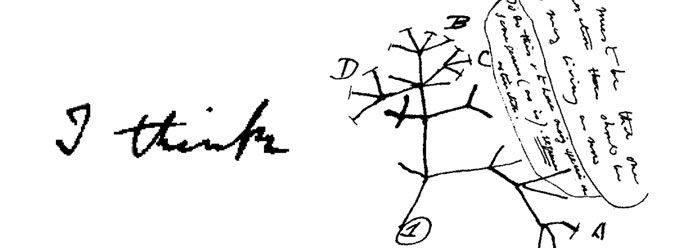
In 1837, Charles Darwin drew his first “evolutionary tree” in his “B” notebook, with the words “I think” scrawled above it, to illustrate his idea that all of today’s species arose from a single common ancestor. But the poor fit of gene sequence data is forcing scientists to abandon the tree.1 “Biologists need to depart from the preconceived notion that all genomes are related by a single bifurcating tree,”2 making way for a new paradigm of origins. A non-Darwinian evolutionary view has been offered, but this proposition is actually just the old “Hopeful Monster” with a twist.3
Darwin’s tree illustrated a long macroevolutionary past that never happened. Darwinian gradualism would predict that over time different species should show a series of slight, successive modifications of genes from simple to complex. However, the tree of life “lies in tatters, torn to pieces by an onslaught of negative evidence…. [D]ifferent genes told contradictory evolutionary stories.”4 Finally, evolutionary biologists seem to be catching up with creation biologists.5
Michael Syvanen, co-editor of Horizontal Gene Transfer (1998) and a medical biochemist at the University of California at Davis, told New Scientist, “We’ve just annihilated the tree of life. It’s not a tree any more, it’s a different topology entirely.”4 For example, one geneticist at the University of Texas at Arlington has found a nearly identical family of DNA “in widely divergent tetrapods.” The same sequence is found in certain animals willy nilly, and is thus “incompatible with [classic Darwinian] vertical inheritance.”6 But without vertical change onward and upward, can the new crazy-bush-like, cross-branching DNA sequence comparisons point to any kind of evolution at all?
French researcher Eric Bapteste and biologist Ford Doolittle in Canada “are at pains to stress that downgrading the tree of life doesn't mean that the theory of evolution is wrong—just that evolution is not as tidy as we would like to believe.”4 The data show that gene distribution patterns are so untidy that neo-Darwinian evolution cannot explain them. Thus, a new mechanism has been suggested.
Donald Williamson, a retired marine biologist, proposed that, at least in starfish, new kinds formed “by the random fusion of two separate species, with one of the partners assuming the role of the larva and the other that of the adult.”4 But how can non-breeding, separate kinds “fuse,” when the biochemistry of reproduction is so highly specified for each kind that “inter-kind breeding” has never been observed? He suggested that “once in a million years it works: the sperm of one species fertilises another and two species become one.”4 Thus, a new version of the Hopeful Monster theory—whereby new kinds are generated not by mutational singularities, but by reproductive singularities—adds to the list of possible mechanisms for evolution.5
Though the patchwork array of genes and other conserved DNA sequences across widely variant species does not fit with the Darwinian evolution model of origins, it does fit an operational structure called “facilitated variation.” This holds that whole genetic modules corresponding to core biological functions can be mixed and matched to varying degrees between generations, within a breeding kind.7 This would allow organisms to adapt to changing environments according to specifically created parameters, but it would not produce the new genetic information that would be needed for upward evolutionary development. Thus, horizontal gene transfer and facilitated variation, as they occur only within breeding kinds, are consistent with both the creation model and the data.
There is no natural law that generates new, specified order (information). Genomes fall apart, but they have never been observed to fall together. The mosaic patchwork of genes found across the living world must, by virtue of sound inference from scientific data, have had its inception in the mind of an all-wise Creator.
References
- In one study, the gene sequence data’s fit to the Darwinian model was 99 percent off: Ciccarelli, F. D. et al. 2006. Toward Automatic Reconstruction of a Highly Resolved Tree of Life. Science. 311 (5765): 1283-1287.
- Dagan, T. and W. Martin. 2006. The tree of one percent. Genome Biology. 7: 118.
- In 1940, Richard Goldschmidt published The Material Basis of Evolution, wherein he revealed that that neo-Darwinian gradual change was an insufficient cause for the incredibly wide variety of today’s living forms. He proposed that every once in a while wild mutational leaps take place, whereby a fundamentally different form—a “hopeful monster”—emerges from what had been a consistently breeding species. His concept has been caricatured by mainstream Darwinists since.
- Lawton, G. 2009. Why Darwin Was Wrong About the Tree of Life. New Scientist. 2692: 34-39.
- “So, we have another new theory to add to the list of hopefuls: Lamarckian evolution; neo-Lamarckian evolution; Darwinian gradualism, neo-Darwinian evolution; punctuated equilibrium; Wright’s shifting balance theory of evolution; theistic evolution; orthogenic evolution among others, and now reticulate evolution. [And now neo-hopeful monster, or random fusion theory.] Which one of these [ten] represents ‘The fact of evolution’? If we put the biological species test to reticulate evolution, we believe it establishes that the hybrid finches and their parents can be nothing more than varieties of a single polymorphic species.” Cumming, K. B. 1997. Reticulate Evolution. Acts & Facts. 26 (7).
- Pace II, J. K. et al. 2008. Repeated horizontal transfer of a DNA transposon in mammals and other tetrapods. Proceedings of the National Academy of Sciences. 105 (44): 17023-17028.
- Thomas, B. What Spurs Evolution: the Old or the Young? ICR News. Posted on icr.org October 17, 2008.
* Mr. Thomas is Science Writer.
Article posted on February 3, 2009.








